酵母是生物大分子药物最常用的表达系统之一。如酿酒酵母、毕赤酵母和汉逊酵母。根据FDA、ICH、CDE等机构规定,和《中国药典》,USP法规要求,满足和药企需求,填补市场空白,赛唐生物开发三种酵母专属HCP ELISA检测试剂盒。
背景:
酵母表达系统是一种最常见的真菌表达系统。通常用于传统疫苗,如疫苗(如乙肝、HPV)、胰岛素类、抗体片段(如VHH);糖蛋白(如EPO)等,相对原核和哺乳动物表达细胞有特殊的优势。如,可进行翻译后修饰,如糖基化,易基因修饰,低免疫原性,低生产成本等,见表1.1。
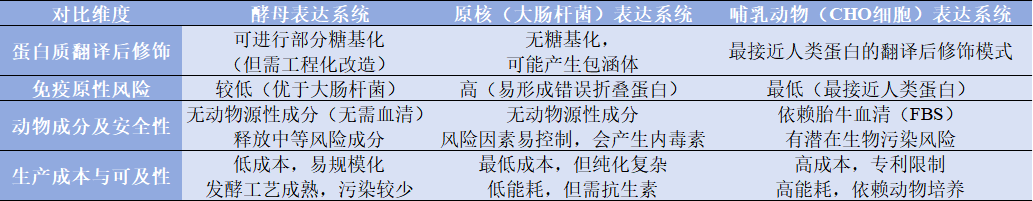
表1.1
酵母对蛋白的糖基化修饰效果较好,又可以分为甲基营养型酵母和非甲基营养型酵母[2]。其中,非甲基营养型酵母,包括酿酒酵母(S288c、A634A、BY4716、CEN.PK、∑1278b、SK1、BJ5464、BY4742 和 W303等)、解脂耶氏酵母(W29、E150、E129、YB423 和 CX161-1B等)。甲基营养型酵母,包括毕赤酵母(Y-11430/X-33 野生型、Mut+ (AOX1+, AOX2+)/Muts (AOX1−, AOX2+)/Mut− (AOX1−, AOX2)甲醇利用不同型、SMD1163 strain (his4pep4prb1)/SMD1165 strain (his4prb1)/SMD1168 strain (his4pep4)蛋白酶活性缺失型、GS115 (his4)/KM71 (Δaox1::SARG4 his4 arg4)/SMD1168 (His4, pep4)组氨酸脱氢酶缺失型)、汉逊酵母(CBS4732 strain (CCY38-22-2,ATCC34438,NRRL-Y-5445) /DL-1 strain (NRRL-Y-7560; ATCC26012)/NCYC495 strain(CBS1976;ATAA14754, NRLLY-1798))[1-3]。
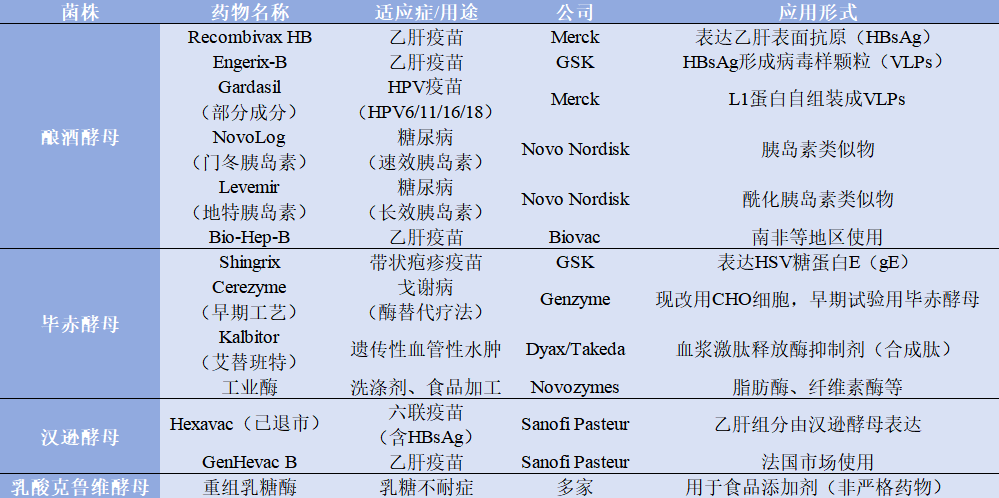
表1.2
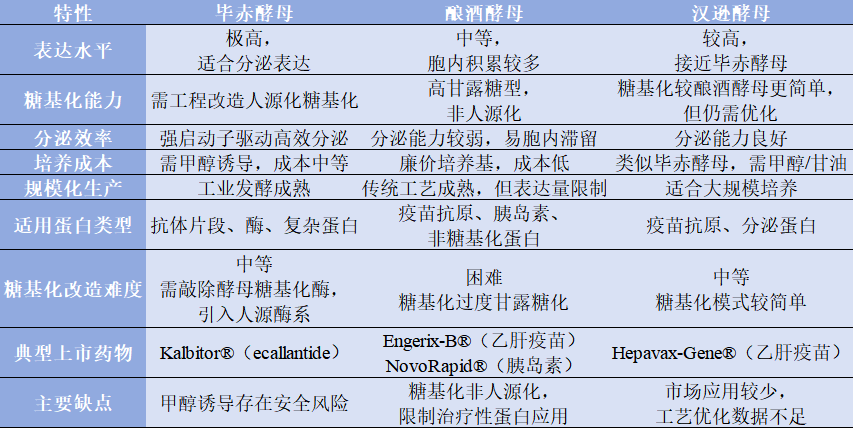
表1.3
1.1毕赤酵母表达系统:
甲基营养型酵母是一种优秀的用于生物医药和工业酶生产的蛋白表达系统。有着细胞高密度培养、容易使用、快速表达、低成本和翻译后修饰等优点。
以毕赤酵母为例,该宿主有着甲醇调节的乙醇氧化酶启动子(PAOX1),高效的分泌构造,转录后修饰和特定培养基中的高密度生长能力[4]。AOX1和AOX2基因编码的乙醇氧化酶,是甲醇氧化通路中的一个酶。根据其突变类型,可以继续分出多种类型菌株系。由于该品系酵母含有AOXI启动子,来代谢甲醇,作为诱导蛋白表达的诱导物。在大批量生产优化后,可以达到20g/L的酶产量。
常用的商业化的毕赤酵母品系有GS115, X-33,Pichia PinkTM和KM71。还有蛋白酶缺陷品系,如SMD1168(his4,pep4,::URA3,ura3)和SMD1168H (pep4);野生型,如BG10,X-33;糖基化品系,如SuperMan5(HIS4+,Och1-disruption), SuperMan5(HIS4+, pep4, Och1 disruption);营养缺陷型品系,如PichiaPink™(ade2), GS115(his4)。配套使用的表达载体有 pJAN-s1(BioGrammatics), pPICZ(Thermo Fisher Scientific), pD902/pD905(DNA2.0)。生产的工业酶,有 Alkaline Xylanase(GS115)、Neutral protease I(GS115)、α -Amylase(GS115)、Lipase(GS115)、 phytase(KM71)、Laccase(GS115)、β-glucosidase(GS115)、 Trypsin(GS115)等。
但是酵母表达药用蛋白的显著突破,是源于FDA许可的Jetrea®和 Kalbitor®。除此之外,还有Human Insulin(X-33), IgG (ScFv fragment)(X-33), HBsAg(GS115),Recombinant Human Interleukin-6(X-33), Human Parathyroid Hormone(Methylotrophic Pichia strain), Recombinant Human Erythropoietin (rhEPO)(X-33)等[5-7]。
来源于非甲基营养型的酿酒酵母表达系统,虽然已经有部分重组蛋白药物上市,但是酵母细胞表现出的蛋白过度糖基化、低产量和胞内质粒不稳定等缺点,限制了其应用。所以也会引入CRIPR/Cas9技术,对特定位点进行突变。如在GlycodExpress™技术专利中介绍,通过连续去除酿酒酵母的甘露糖转移酶(MNN1)和糖基转移酶基因,增强了N-糖基化同质性,并增加了高尔基体细胞器中的UDP-GlcNAc的转运能力[8]。
1.2酿酒酵母表达系统:
以啤酒酵母为例,其基因组是1996年第一个被完全测序完成的真核基因组。在工业生产乙醇方面,啤酒酵母细胞,有着pH范围宽,乙醇和糖浓度高,渗透压强等众多特点,优于细菌、其他酵母和丝状真菌[9]。同时也是最常用的基因表达调节模型,且在信号转导,衰老,凋亡,代谢,细胞周期控制,细胞程序性死亡,神经退行性疾病,自噬,分泌路径等重要生物学过程。啤酒酵母生产的异源糖蛋白,会导致异常的过度甘露糖基化,并会导致蛋白活性降低和免疫原性增强。该甘露糖基化修饰的通路,主要与Mnn2p和Mnn11p基因有关。N-糖基化修饰导致蛋白分泌增加[10]。敲除α-1,6-甘露糖转移酶Och1p,能够生产出有活性形式的人组织型纤溶酶原激活剂。
1.3汉逊酵母表达系统:
汉逊酵母,即多形汉逊酵母。是一种特殊的甲醇营养型酵母,其构建的系统,可利用强诱导型的启动子如FMD, MOX, GAP等。表达的异源蛋白可实现糖基化修饰,可利用完全合成培养基进行高密度发酵,其遗传背景较为清楚,安全又高效的性能使得近年来被越来越多的研究人员作为最有前景的真核表达系统。
汉逊酵母有两个有效的表达元件:1.启动外源基因高效表达的载体系统;2.用于筛选标记的特殊基因。其筛选标记含两类:一、营养缺陷型标记,如URA3、LEU2和HIS4等发生缺失条件无法生长的宿主菌标志基因;二、是显性选择标记,如抗G418、抗Zeocin基因等。一般基因缺陷型酵母菌的制备,都是通过化学诱变、杂交或原生质体融合等,来获得对应的标记基因。
总之,酵母表达系统是一个物美价廉的药物表达宿主。对于免疫方面,如乙肝疫苗(Engerix-B®)、HPV疫苗(Gardasil®),酵母表达的疫苗有更强的免疫原性,稳定的VLPs结构。
在有相同疗效的情况下,酵母表达药物的成本更低,如门冬胰岛素(NovoRapid®);对于抗体片段类药物,Such as Obinutuzumab(Gazyva®)、Caplacizumab(Cablivi®)ADCC更强(糖基化优化),组织穿透性更好;表达细胞因子Epoetin delta(Dynepo®),也有着更长的半衰期。
综上,毕赤酵母、酿酒酵母和汉逊酵母在亚洲区域的药企的应用较为常见,三种酵母在表达水平,分泌效率,糖基化能力及其改造难度,基因修饰能力,适合药物类型的能特点上,均有较大差异。其各自优缺点比较,和上市药物表达宿主酵母类型,见表1.2、表1.3。
数据:
2.1标准曲线:
上海赛唐生物药物残留相关产品:酿酒酵母HCP,毕赤酵母X-33 HCP,汉逊酵母HCP ELISA Kit,其4PL拟合R²均达到0.999,见图一。
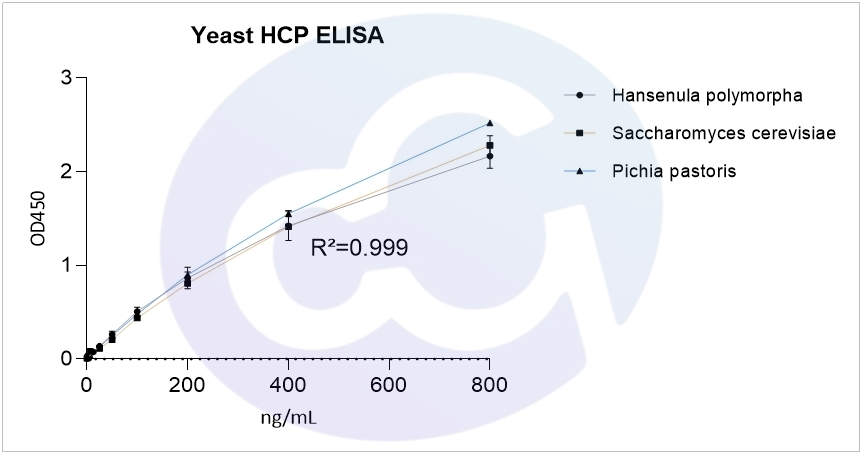
图一
2.2 质控数据:
毕赤酵母X-33,汉逊酵母和酿酒酵母HCP标曲调试在0-800ng/mL范围。测定最低检测限和最低定量限。并测试三种酵母HCP的交叉反应,和精密度等。另外在特定浓度尿素,盐浓度和pH条件下,回收率均有优异效果,见表2.1。
.png)
表2.1
参考标准:
ICH Q6B: Specifications: Test Procedures and Acceptance Criteria for Biotechnological/Biological Products.
ICH Q7: Good Manufacturing Practice Guide for Active Pharmaceutical Ingredients.
ICH S6 (R1).
EMA: Guideline on Immunogenicity assessment of therapeutic proteins (2017).
FDA: Points to Consider in the Manufacture and Testing of Monoclonal Antibody Products (1997).
参考文献:
[1] Fickers, P. Pichia pastoris: A workhorse for recombinant protein production[J]. Current Research in Microbiology and Biotechnology, 2014, 2(3):354-363.
[2] Vanz, A., Lünsdorf, H., Adnan, A., et al. Physiological response of Pichia pastoris GS115 to methanol-induced high level production of the Hepatitis B surface antigen: Catabolic adaptation, stress responses, and autophagic processes[J]. Microbial Cell Factories, 2012, 11(103):1-11.
[3] Stöckmann, C., Scheidle, M., Dittrich, B., et al. Process development in Hansenula polymorpha and Arxula adeninivorans, a reassessment[J]. Microbial Cell Factories, 2009, 8(22):1-10.
[4] Vieira, S. M., da Rocha, S. L. G., da Neves-Ferreira, A. G., et al. Heterologous expression of the anti myotoxic protein DM64 in Pichia pastoris[J]. PLoS Neglected Tropical Diseases, 2017,11(7):1-20.
[5] Meehl MA, Stadheim TA. Biopharmaceutical discovery and production in yeast[J]. Current opinion in biotechnology. 2014, 30:120-127.
[6] Thompson CA. FDA approves kallikrein inhibitor to treat hereditary angioedema[S]. American journal of health-system pharmacy: AJHP: official journal of the American Society of Health-System Pharmacists. 2010, 67(2):93.
[7] Imran Safder, Sajad Khan, Iram-us Islam, et al. Pichia pastoris expression system: a potential candidate to express protein in industrial and biopharmaceutical domains[J]. Biomedical Letters. 2018. 4(1):1-14.
[8] Piirainen, M. A., Boer, H., de Ruijter, J. C., et al. A dual approach for improving homogeneity of a human-type N-glycan structure in Saccharomyces cerevisiae[J]. Glycoconjugate Journal, 2016, 33(2):189-199.
[9] Tesfaw, A., & Assefa, F. Current trends in bioethanol production by Saccharomyces cerevisiae: Substrate, inhibitor reduction, growth variables, coculture, and immobilization[J]. International Scholarly Research Notices, 2014, 2014:1-11.
[10] Tang, H., Wang, S., Wang, J., et al. N-hypermannose glycosylation disruption enhances recombinant protein production by regulating secretory pathway and cell wall integrity in Saccharomyces cerevisiae[J]. Scientific Reports, 2016, 6(1):1-13.
上海赛唐生物技术有限公司,专注于生物医药工业检测领域15年研发,推出HCP残留检测系列产品,以及HCP专属性抗体制备、覆盖度分析等全流程技术服务!
https://www.bluegene.cc/index.asp
产品类型:
| Cellgene Bioscience-Drug Residue Products | |
| Host Cell Protein ELISA kits (HCP) | |
| CH-K0018-2 | CHO Host Cell Protein (CHO HCP) ELISA kit, G2 |
| CH-K0018-3 | CHO Host Cell Protein (CHO HCP) ELISA kit, G3 |
| HH-H0019-2 | HEK293 Host Cell Protein (HEK293 HCP) ELISA kit, G2 |
| EH-E0020-3 | E. coli Host Cell Protein (E. coli HCP) ELISA kit, G3 |
| PH-E0021-3 | Pichia Pastoris Host Cell Protein (PP HCP) ELISA kit, G3 |
| HP-H0023-3 | Hansenula Polymorpha Host Cell Protein ELISA kit, G3 |
| SC-H0024-3 | Saccharomyces Cerevisiae Host Cell Protein ELISA kit, G3 |
| SF-H0025-3 | Spodoptera Frugiperda 9 Host Cell Protein ELISA kit, G3 |
| Medium Residues Detection kits | |
| NEGES0890 | Protein A ELISA Kit (Boiling) |
| NEGEP0890 | Protein A ELISA Kit |
| NE03I0431 | Mouse Immunoglobulin G ELISA Kit |
| NE11I0431 | Bovine Immunoglobulin G ELISA Kit |
| NE01I0431 | Human Immunoglobulin G ELISA Kit |
| NE06I0431 | Goat Immunoglobulin G ELISA Kit |
| NEGEK0006 | Kanamycin ELISA Kit |
| NEGEP1270 | Protein L ELISA Kit |
| NEGEP1271 | Protein G ELISA Kit |
| NEGES0014 | Bovine Serum Albumin ELISA Kit |
| NEGES0015 | Human Serum Albumin ELISA Kit |
| NEGED0018 | Dextran Sulfate Salt Detection kit |
| Host Cell DNA Detection kits (HCD) | |
| NS-D050T/NS-D100T | NS0 Host Cell DNA (NS0 HCD) Residue Detection kit |
| EC-D050T/EC-D100T | E.coli Host Cell DNA (E.coli HCD) Residue Detection kit |
| VE-D050T/VE-D100T | Vero Host Cell DNA (Vero HCD) Residue Detection kit |
| HK-D050T/HK-D100T | HEK293 Host Cell DNA (HEK293 HCD) Residue Detection kit |
| CH-D050T/CH-D100T | CHO Host Cell DNA (CHO HCD) Residue Detection Kit |
| PP-D050T/PP-D100T | Pichia Pastoris Host Cell DNA (PP HCD) Residue Detection Kit |
| CG-DP050/CG-DP100 | Magnetic Residual DNA Sample Preparation Kit |
| Host Cell Protein Antibodies | |
| CH-K0018-3-Ab | CHO Host Cell Protein G3 Antibody |
| CH-K0018-2-Ab | CHO Host Cell Protein G2 Antibody |
| EH-E0020-3-Ab | E.coli Host Cell Protein G3 Antibody |
| PH-E0021-2-Ab | Pichia Yeast Host Cell Protein G2 Antibody |
| HH-H0019-2-Ab | HEK293 Host Cell Protein G2 Antibody |
| Buffer Products | |
| CG-H0100 | HCP ELISA buffer |
| CG-P0200 | Protein L ELISA buffer |

VX:13764544806
WEB:www.bluegene.cc
E-mail:sales@bluegene.cc
TEL:400-882-6373 / 021-61106433



